微生物の特異な代謝機構に関する研究
メタン発酵系微生物の共生的プロピオン酸酸化メカニズムの解明
限られたエネルギーを最大限に利用するメカニズムを知る
メタン発酵条件下では酸素、さらには硫酸や硝酸、鉄イオンといった電子受容体がほとんど存在しない。しかしながら、この環境に最適化している微生物は有機物に残る還元力を利用し生育する。この還元力利用のメカニズムにはこの環境に生育する微生物特有のものがあり、それを知ることが限られたエネルギーを利用する生物特有の新たなメカニズムにつながると考えている。
このメタン発酵を利用するメタン発酵槽では、有機酸の蓄積により発酵槽が不安定になることがしばしば見られる。その蓄積の原因として有機酸の分解に異なる微生物の協調、時には共生が必要であることが挙げられる。この協調にはそれぞれの微生物の持つ限られたエネルギーを利用する機構が大きく関係している。
この研究では、種の異なる微生物であるプロピオン酸酸化細菌 Pelotomaculum thermopropionicum SIと水素資化性メタン生成アーキア Methanothermobacter sp. CaT2 が特定の状態でお互いが生育するために共生する過程に着目し、この2つの菌の共生機構を明らかにする事で、2つの異なる生命システムが協調及び反発する原理や融合して新たな生命システムを形成するメカニズムを理解する事を目的としている。
嫌気的なプロピオン酸酸化は微生物共生により進行する
メタン発酵槽で蓄積しやすい有機酸であり、低級揮発性脂肪酸の一つであるプロピオン酸の酢酸への酸化反応は、標準状態では吸エルゴン反応であり自発的には進行しにくい。つまり、プロピオン酸酸化の結果生成される水素(もしくはギ酸)が非常に低濃度で維持されないと、継続的に微生物が生育する事が出来ない。そこで、生成される水素をメタン菌生成菌が資化しメタンにすることで、水素濃度が低く保たれることになる。このように、2つの異なる微生物は共生することでプロヒオン酸からメタンを生成し、お互いにエネルギーを獲得している。

我々はこのプロピオン酸からメタン生成を行う共生系が、2つの異なる種が共生する事によって創発的な機能発現を行うモデルとなりうると考えている。そこで、まずそれぞれの微生物がどのようにプロピオン酸からメタンを生成しているのか、その代謝及び機構を明らかにしようとしている。さらに、この共生において重要な役割を果たすと考えている凝集の形成過程とその形成に関与する分子に関する知見を取得している。
嫌気的プロピオン酸酸化はメチルマロニルCoA経路が主に用いられる
電子受容体のない状態での嫌気的なプロピオン酸の酸化は、メチルマロニルCoA経路(MMC pathway)により進行する。細胞内に取り込まれたプロピオン酸は、メチルマロニルCoAを中間代謝物として、コハク酸、フマル酸、オキサロ酢酸、ピルビン酸を経て酢酸へと酸化される(下図)。
このようなプロピオン酸酸化経路は、中温(37℃付近)菌での解析は Syntrophobacter fumaroxydance の代謝解析やゲノム解析を基に行われているが(Plugge et al., 2012)、我々は高温(55℃付近)で生育する P. thermopropionicum のゲノム解析や発現タンパク質等の解析を実施し、この経路が実際に P. thermopropionicum で利用されていることを予測した(Kosaka et al., 2006)。さらには全ゲノム配列を解析し、P. thermopropionicum の遺伝情報を明らかにした(Kosaka et al., 2008)。これらの解析から、共生が可能なプロピオン酸酸化細菌はこの環境に適応してゲノムを進化させてきたことが示唆された。現在は mmc pathway の詳細な解析を行っている。
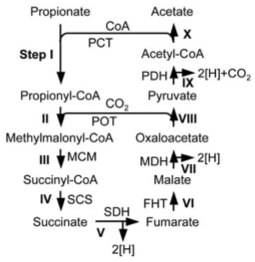
メチルマロニルCoA経路
微生物の共生では凝集がよくみられる
異なる微生物が凝集することによりプロピオン酸代謝効率の向上が示唆されている(Ishii et al., 2005)。我々は、その凝集が可能な高温性水素資化性メタン生成菌 Methanothermobacter sp. CaT2を分離し、ゲノム配列を報告した(Kosaka et al., 2013)。このメタン生成菌は表層糖を持ち、自己凝集を形成する(Kosaka et al., 2014)。この凝集に関係する細胞の因子の分析を試みたところ、変異株の取得によって表層に局在するタンパク質がこのメタン生成菌の凝集に大きく関与することを示唆した。またその凝集にはMg2+やCa2+といった二価の金属イオンが関与することも示唆した(Sumikawa et al., 2019)。一方、凝集性が強化された変異株でタンパク質以外の因子がその強化に関与することを示唆した(Sumikawa et al., 2020)。